Research Focus
1) Signal evolution: My past research work focused on understanding the impact of some drivers of signal evolution including habitat structure and heterospecific masking using crickets and katydid of a rainforest as a model system. My current research interests continue to lie in understanding the selective forces that shape acoustic signals and influence signalling behaviour. However, the factors that I am focusing on currently include a) Sociality b) Masking avoidance and c) Sexual selection and we are examining this in both invertebrate and vertebrate model systems.
2) Impact of anthropogenic activities on sensory ecology: Humans have left their footprints on various aspects of the biology of non-human animals. Their activities have significantly impacted the sensory ecology of animals. Understanding this impact and how animals deal with these changes is another major focus of my current research. Towards this we are examining the impact of artificial light and traffic noise on signalling behaviour in animals. In addition we have also examined the negative impact of turbidity on fish and the behavioural plasticity they exhibit to deal with this problem.
3) Natural history and life history: My lab, since its inception, included people who loved to observe animals and from that shared primordial interest stemmed out appreciation of natural history and urge to understand life histories of animals. Towards the former we have been involved in taxonomic studies on insects (crickets and moths) using classical taxonomical and molecular tools. We have also been studying the activity and infection patterns in birds and insects at different temporal scales. Further, using insects as a model system, we examine the effect of rising temperatures on organisms.
We are also actively involved in monitoring species diversity and distribution across India.
Towards all of this we are using bioacoustics as a tool. The choice of the model system depends on logistical considerations and the question asked. As far as possible we adopt non-invasive methods for our observations and experiments to address questions using commonly available species that are widely distributed. Some description of ongoing projects under each of these areas are given below for the benefit of the general public, future students, potential collaborators and funding agencies. If you find any of these interesting, do drop us a line over email. We look forward to hearing from you!
Field sites
1) Western Ghats 2) North Western Himalayas 3) Terai grasslands and forests 4) Andaman Nicobar islands 5) Central Indian Highlands
Why are signals so structured? Which selective forces shape signals? What information is encoded in signals?

What role does sociality have in the evolution of complex acoustic communication? Do social animals have higher communicative complexity? What are the features of complex acoustic signals?
We are examining these and other questions as part of a long-term project on social babbler found in India. We are working on various aspects of the biology and behaviour of different babblers and we are keen to collaborate with interested researchers. Read about our findings here.

How do animals communicate effectively in noisy choruses? Do signalers with similar acoustic features avoid calling together? Do crowded acoustic communities exhibit overdispersion of acoustic traits? How do conspecifics avoid being masked by each other, given overall signal similarity is high?
I examined heterospecific masking avoidance and later with my student studied conspecific masking avoidance in crickets. We find that while the problem is common, the solutions are very different. Read more here and here.
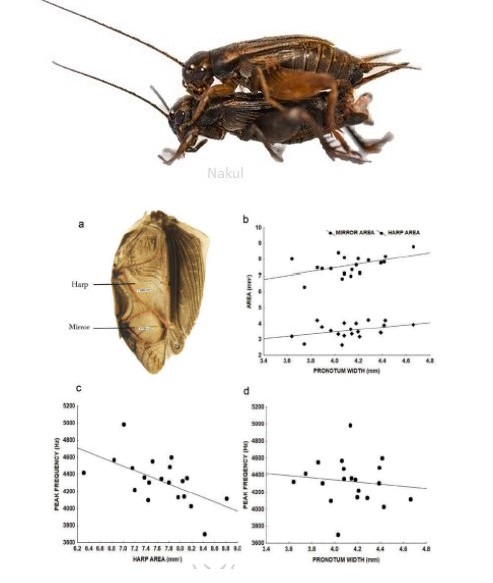
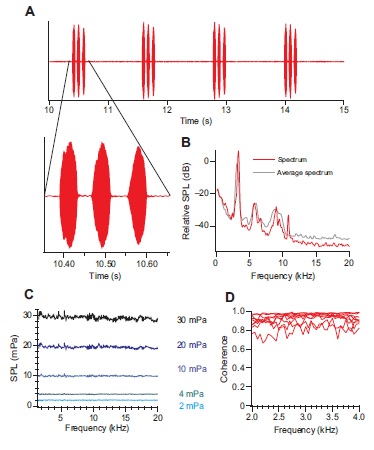
Why do crickets produce postcopulatory calls? What information is encoded in the mating and the aggressive call? Do receivers use this information in their decision making?
We are examining these questions in the context of both male-male competition and female choice in field crickets. One of our findings suggests that the peak frequency of the mating calls indicates male body size. Read more here.
How have human activities altered the sensory environment of animals? How do animals cope with it?

How does turbidity impact visually orienting animals? Can acclimatization mitigate the negative impact? How do fish find foraging sites when their visual environment is altered?
We used Zebrafish to find out. Turbidity severely impacted foraging efficiency in zebrafish, but acclimatization mitigated the impact at least partially. We also show that fish can utilize conspicuous and familiar visual landmarks under turbid condition to navigate effectively!
Read about our findings here.

S
ince the advent of the light bulb, the nocturnal landscape has been altered significantly. The impact of Artificial Light at Night (ALAN) has been studied extensively on diurnal animals that must rest at night. But what about nocturnal animals who evolved to be active at night?
We are studying this using crickets and they seem to be impacted significantly. We are investigating measures that policy makers may adopt to address this. Meanwhile, let's hope that the 'dark night rises again'.

One of the most pervasive impact of rapid urbanization is the increase in environmental noise, especially from traffic. How have the urban soundscapes been altered by traffic noise? What impact does anthropogenic noise have on acoustically communicating animals? How do animals cope with this sensory pollution?
Given that we live and work in an Indian city, we are perfectly placed to study these. We are addressing these questions using both diurnal and nocturnal animals to ask if we drowning out nature's sounds?

Reliable identification of species rests on rigorous taxonomic work which is time and effort intensive. It requires specialized skills and such trained taxonomists are hard to find. Not enough people are being trained or retained in this important field.
At BEL we value the importance of this field and in our modest way continue to contribute to it. We have worked on cricket and moth taxonomy. We have reported over 70 moth species as new records from Punjab. Moreover, we have describe a new species of cricket from India belonging to the genus Teleogryllus. Read moe about it here.
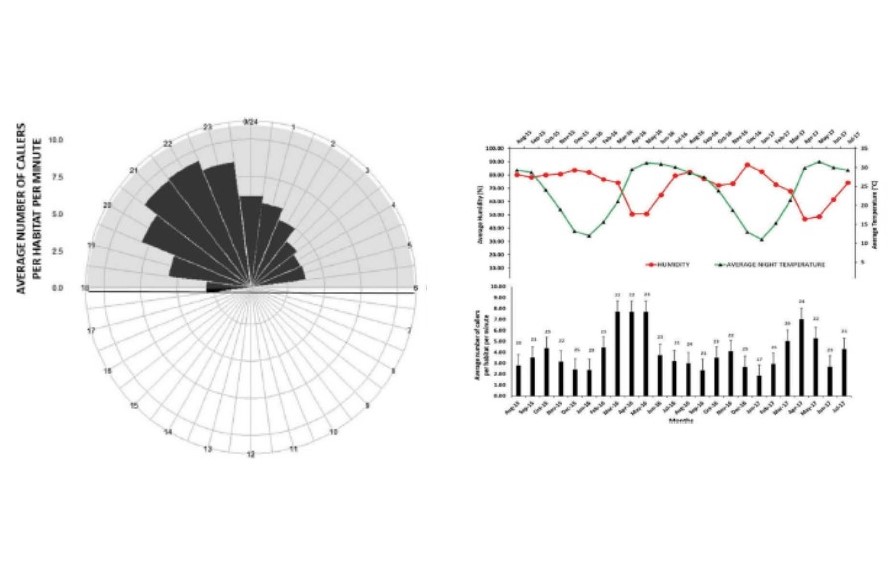
Environmental conditions across diel and seasonal scale shape how animals organize their behaviour across temporal scale. Understanding these activity patterns and the factors that drive them are crucial aspects of organismal biology.
We have been studying the diel and seasonal patterns of activities in both diurnal and nocturnal animals, including birds, crickets, moths and fruit bats. This always involves rigorous natural history observations in the field, often at ungodly hours. However, the treasure of information such studies generate make everything worth the while. Read about some of our findings here and here.

Impact of rising temperatures
Does increased ambient temperature impact the growth, development and behaviour of insects?
We examined the effect of rearing temperature on growth and development of crickets and on the mating calls of adult males. We find that crickets reared in warmer temperatures are bigger and exhibit faster development. We also find that the mating calls of crickets reared in warmer temperature are different from those reared in cooler temperature. This is in addition to the impact of the immediate ambient temperatures on cricket calls! Our findings have implications on the effect of climate change on insects. Read the paper here.
The one process now going on that will take millions of years to correct is the loss of genetic and species diversity by the destruction of natural habitats. This is the folly our descendants are least likely to forgive us.
We have been working towards biodiversity monitoring and education using acoustics.
Bats: I was involved in developing the first echolocation call library of bats of India. We showed that the echolocation call serve as a reliable indicator of the species identity. This can now be used for passive acoustic monitoring of bats. Read more here.
Cricket: We have been conducting surveys for cricket species diversity across India including North-western Himalayas and the foothills, Terai grasslands and forests in Dudhwa National Park, the caves of Chattishgarh, Central India and Western ghats. Whenever possible we also record their calls and aim to build a mega repository of cricket calls from across India.
Moths: We have been monitoring moth diversity of Shivaliks and have developed a field guidebook for young naturalists to help in quick identification of moths.
Birds: In collaboration with Jimmy Bahuleyan and three undergrads from IISER Mohali we have developed an Android app for bird identification on various IISER campuses. The app has birds of IISER Mohali and IISER Bhopal. Several other IISER campuses will be added shortly. We are working on it to continuously update it and add more features. Interested students are requested to drop in a line.
Background pic by Mahandran V.

Mahandran V, Hakkim H, Sinha V and Jain M. (2022). Fruit scent as an indicator of ripeness status in ‘bat fruits’ to attract ‘fruit bats’: chemical basis of chiropterochory. Acta Ethologica, 1-11.
Yambem S. D. and Jain M. (2022). Temporal variation in the behaviour of a cooperatively breeding bird, Jungle Babbler (Argya striata). Tropical Ecology, 1-13.
Jaiswara R., Desutter-Grandcolas L. and Jain M. (2021). Taxonomic revision of Teleogryllus mitratus (Burmeister, 1838) and T. occipitalis (Serville, 1838) in India, with the description of Teleogryllus rohinae Jaiswara amp; Jain sp. nov. and a key for Teleogryllus species from India (Orthoptera: Gryllidae). Zootaxa, 5016(1): 81-106.
Yambem S. D., Chorol S. and Jain M. (2021). More than just babble: functional and structural complexity of vocalizations of Jungle Babbler. Behavioral Ecology and Sociobiology 75 (8), 1-14
Singh R. and Jain M. (2021). Variation in call types, calling activity patterns and relationship between call frequency and body size in a field cricket, Acanthogryllus asiaticus. Bioacoustics, 1-19.
Singh R., Prathibha P. and Jain M. (2020). Effect of temperature on life-history traits and mating calls of a field cricket, Acanthogryllus asiaticus. Journal of Thermal Biology, 93: 102740
Sekhar M., Singh R., Bhat A. and Jain M. (2019). Feeding in murky waters: acclimatization and landmarks improve foraging efficiency of zebrafish (Danio rerio) in turbid waters. Biology Letters, 15(7), 20190289.
Jain M. (2018). Amotz Zahavi: Tale of a swamp and peacock tails. Resonance, 23: 525-533.
Raghuram H., Jain M. and Balakrishnan R. (2014). Species and acoustic diversity of bats in a palaeotropical wet evergreen forest in southern India. Current Science, 107(04):631-641.
Balakrishnan R., Bahuleyan J., Nandi D. and Jain M. (2014). Modeling the effects of chorus species composition and caller density on acoustic masking interference in multispecies choruses of crickets and katydids Ecological Informatics, 21: 50-58.
Jain M., Diwakar S., Bahuleyan J., Deb R. and Balakrishnan R. (2014). A Rain Forest Dusk Chorus: cacophony or sounds of silence? Evolutionary Ecology, 28(1): 1-22.
Rajaraman K., Mhatre N., Jain M., Postles M., Balakrishnan R. and Robert D. (2013). Low pass filters and differential tympanal tuning in a paleotropical bushcricket with an unusually low frequency call. Journal of Experimental Biology, 216: 777-787.
Jain M., Balakrishnan R. (2013). From leaf litter to canopy: noninvasive and reliable sampling in a tropical rainforest. In Treetops at Risk. Challenges of Global Forest Canopies (Eds) Lowman M., Soubadra M. D. and Ganesh T. Springer Verlag.
Jain M. and Balakrishnan R. (2012). Does acoustic adaptation drive vertical stratification? A test in a tropical cricket assemblage. Behavioral Ecology, 23(2): 343-354.
Jain M. (2012). Songsters in the canopy: cricket acoustics. In Forest Canopies of South Asia- a glimpse (Eds) Soubadra M. D., Ganesh T. and Tripathy A. ATREE.
Jain M. and Balakrishnan R. (2011). Microhabitat selection in an assemblage of crickets (Orthoptera: Ensifera) of a tropical evergreen forest in Southern India. Insect Conservation and Diversity, 4(2): 152-158.
Jain M., Kuriakose G. and Balakrishnan R. (2010). Evaluation of methods to estimate foliage density in the understorey of a tropical evergreen forest. Current Science, 98(4): 508-515.
Diwakar S., Jain M. and Balakrishnan R. (2007). Psychoacoustic sampling as a reliable, non-invasive method to monitor orthopteran species diversity in tropical forests. Biodiversity and Conservation, 16 (14): 4081-4093.